<sup id="kwqog"></sup>
噬菌體能展示各種分子的功能域或抗原、抗體的相應(yīng)配體結(jié)構(gòu)
方法:將目標(biāo)噬菌體感染大腸桿菌ER2537進(jìn)行擴(kuò)增
該病毒無(wú)囊膜
病原學(xué)檢查首先采用糞檢,可用直接涂片法
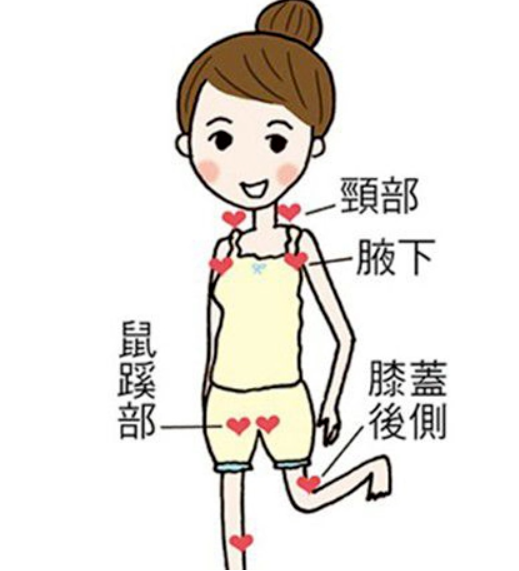
①糞便檢查:直接涂片檢查蟲卵,毛蚴孵化法等
本文地址:http://www.mcys1996.com/jiankang/154943.html.
聲明: 我們致力于保護(hù)作者版權(quán),注重分享
Copyright ? 2023-2024 大道家園 版權(quán)所有
備案號(hào): 陜ICP備2022010374號(hào)-1
聯(lián)系方式:douchuanxin@foxmail.com


.png)
![]()
身體不適時(shí)請(qǐng)至正規(guī)醫(yī)院就診
2020国产精品久久精品|
欧美日韩视频在线第一区|
日韩A无V码在线播放|
久久精品国产导航|
精品无码国模私拍视频|
日本婬乱A片AAA毛片麻豆软件|
大陆国语对白国产av片|
国产又黄又爽又刺激的免费网址|
国模吧无码一区二区三区|
日韩精品一区二区午夜成人版|
新版天堂资源中文8在线|
人妻av无码av中文av日韩av|
国产精品午夜自在在线|
国产香蕉尹人在线视频你懂的
|
国产精品全国免费观看高清|
亚洲欧洲美洲无码精品Va|
97色精品视频在线观看|
亚洲综合av色婷婷|
成年女人免费碰碰视频|
亚洲人成无码网站在线观看|
97精品人妻一区二区三区香蕉|
国产一区二区怡红院|
国产97成人亚洲综合在线观看|
色五月丁香五月综合五月4438|
国产精品 精品国内自产拍|
狠狠做五月四房深爱婷婷|
成人综合色在线一区二区|
国产成人亚洲精品|
久久国产午夜理论片|
性一交一乱一乱一视频|
AV无码A片高潮AV|
国产av寂寞骚妇|
国产精品对白刺激久久久|
18禁黄网站免费|
亚洲精品无码久久久久av老牛|
国产亚洲精品aa片在线观看网站|
无码少妇一区二区三区视频|
亚洲精品在看在线观看|
内射人妻无套中出无码|
国产午夜精品一二区理论影院|
欧美精品一国产成人综合久久|
东京热加勒比无码视频|
免费看999永久A片视频|
…中文天堂最新版在线网|
国产午夜人做人免费视频|
刺激性A片欧美激情免费|
日本一区二区三区免费不卞|
亚洲欧美闷骚少妇影院|
久久精品亚洲精品无码|
华人少妇被黑人粗大的猛烈进
|
а√资源新版在线天堂|
少妇高清精品毛片在线视频|
最近更新中文字幕免费版|
少女视频成全观看免费完整观看|
国产一起色一起爱|
国产在线aaa片一区二区99|
HD欧美free性XXXX护士|
亚洲一区二区三区中文字幕在线|
国产精品乱码一区二区三区|
成人性生交大免费看|
爽爽影院免费观看视频|
九九九九精品视频在线观看|
中文字幕亚洲无线码a|
玩弄白嫩少妇xxxxx性|
亚洲在AV无码天堂|
婷婷色国产精品视频一区|
一夲道人妻熟女AV网站|
久久无码人妻国产一区二区|
成AV人片一区二区三区久久|
日韩精品久久久久久久电影蜜臀|
午夜精品久久久久久久爽|
亚洲国产一区二区三区亚瑟|
午夜影视啪啪免费体验区入口
|
国产在线看片免费人成视频|
性xxxx视频播放|
亚洲午夜成人精品无码|
人人妻人人添人人爽欧美一区|
无码人妻丝袜在线视频|
丰满熟妇人妻av无码区|
精品国产第一福利网站|
日产精品卡二卡三卡四卡区满十八|
亚洲精品无码久久毛片波多野吉衣
|
国产99996在线亚洲|
亚洲国产成人精品无码一区二区|
久热这里只有精品视频6|
久久婷婷五月综合国产尤物APP|
欧美人与禽交zozo|
铜铜铜铜铜铜铜铜铜好大好痛|
无码人妻久久一区二区三区免费丨|
中文在线网在线中文|
婷婷五月深深久久精品|
亚洲国产精品久久青草无码|
成人无码精品一区二区三区亚洲区|
一线高清中文免费观看视频
|
五月天社区|
国产欧美VA欧美VA香蕉在线观看|
影音先锋a天堂2017|
天堂中文在线8最新版地址|
国产99视频精品免视看9|
黑寡妇巨大粗爽毛片欧美|
久久天天躁狠狠躁夜夜av浪潮|
亚洲精品午夜久久久伊人
|
乱相姦处破女AV毛片
|
国精产品W灬源码1H855.C|
99久久国产综合精品1|
无码少妇一区二区性色AV|
亚洲妇女无套内射精|
日韩精品无码人妻一区二区三区|
越南少妇BBV叉叉叉|
亚洲丁香五月激情综合|
久久久久无码精品国产h动漫|
人爽人澡人妻a片精品二区|
欧美黑寡妇AAAAA片|
又爽又黄无遮拦成人网站|
爱做久久久久久|
婷婷综合另类小说色区|
和搜子同居的日子av片|
激情五月色综合国产精品|
JLZZJZZ免费大全|
亚洲国产精品成人无码区
|
亚洲精品WWW久久久久久|
中文字幕人妻丝袜美腿乱|
3Dmax亚洲NINEBOXmod|
激情综合色五月丁香六月欧美|
人妻丰满熟妇AV无码区HD|
国产精品丝袜肉丝出水|
日本熟妇XXXXX乱|
精品黑人一区二区三区|
成人亚洲精品久久久久软件|
内射人妻少妇无码一本一道
|
色偷拍 自怕 亚洲 10p|
亚洲精品国产一区二区|
在线观看免费人成视频播放|
亚洲成A人片在线观看无码|
国产精品扒开腿做爽爽爽a片唱戏
无码免费一区二区三区
|
成人综合伊人五月婷久久
|
粉嫩极品美女国产在线观看|
99国精产品灬源|
亚洲国产美女精品久久久久∴|
亚洲熟妇图片[13p]|
午夜裸体性播放|
亚洲中文无码成人片|
五月天激情电影|
夜夜高潮夜夜爽国产伦精品|
久久精品国产99久久美女|
亚洲人成网站在线在线观看|
日韩精品无码成人专区AV
|
精品成人无码a片观看香草视频|
亚洲乱码伦小说区|
成人欧美一区二区三区在线观看|
少妇夜夜春夜夜爽试看视频|
又黄又爽又色的视频|
自拍亚洲一区欧美另类|
真人做爰片免费观看播放第09集|
国模吧无码一区二区三区|
exo妈妈mv高清视频播放|
久久九九久精品国产|
四虎永久在线精品免费观看
|
阿v网站在线观看|
国产+麻豆视频|
国产精品无码久久久久久|
久久亚洲精品中文字幕|
老色鬼永久精品网站|
性夜夜春夜夜爽AA片A|
国产成年女人特黄特色毛片免
|
丁香五月亚洲综合在线|
天堂俺去俺来也WWW色官网|
四虎国产精品免费永久在线|
强奷乱码中文字幕乱老妇|
亚洲精品无码不卡久久久久|
亚洲国产精品无码AAA片|
777777777妇女亚洲|
日韩精品东京热无码视频|
亚洲高清无码视频网站在线|
成熟丰满熟妇高潮xxxxx|
97资源共享在线视频|
中日韩亚洲人成无码网站|
五级黄高潮片90分钟视频|
日本www.在线中文字幕|
少妇系列之白嫩人妻|
国产成人AV在线免播放观看更新|
久久综合亚洲色一区二区三区|
亚洲欧美第一成人网站7777|
人妻无码中文字幕|
成人国产三级在线观看|
午夜精品A片一区二区三区资源看|
人禽无码视频在线观看|
国产成A人亚洲精V品无码|
亚洲国产成人无码专区|
婷婷久久久亚洲欧洲日产国码av|
欧美亚洲偷图色综合|
无码人妻一区二区三区免费手机|
日韩制服国产精品一区|
亚洲熟妇AV一区|
成人免费无码不卡毛片视频|
婷婷色婷婷开心五月四房播播久久|
一本一道波多野结衣AV黑人|
中文成人无码精品久久久不卡|
日本妇人成熟免费中文字幕|
亚洲三级香港三级久久
|
蜜臀AV无码国产精品色午夜麻豆
|
亚洲午夜精品一区二区|
大地资源免费更新在线播放|
成片免费观看视频在线|
欧美性狂猛xxxxxbbbbb|
熟女体下毛毛黑森林|
真人做爰片免费观看播放第09集|
扒开老师双腿猛进入白浆小说|
五月丁香综合缴情六月小说|
av无码国产精品午夜a片|
欧美重口变态另类A片|
成熟女人特级毛片www免费|
大色堂午夜福利国产TV6080|
内射合集对白在线|
一区二区三区无码按摩精油|
国产人久久人人人人爽|
在线精品亚洲一区二区|
亚洲欧洲精品专线|
精品无码国产一区二区三区麻豆
|
亚洲日韩乱码中文无码蜜桃|
无码精品视频一区二区三区|
欧美日韩视频在线第一区|
丝袜无码一区二区三区|
最近免费中文字幕大全免费|
五月天中文字幕mv在线|
日韩在线一区二区不卡视频|
亚洲欧洲∨国产一区二区三区|
国精产品69永久中国有限|
夜夜躁日日躁狠狠久久av
|
欧美日韩视频在线第一区|
69xx老熟女|
97久久超碰成人精品网页|
亚洲性线免费观看视频成熟|
午夜两性刺激视频免费看|
精品欧美小视频在线观看|
国产九九久久99精品影院|
国产情侣A片A毛片手机在线|
久久婷婷五月综合色国产免费观看
|
久久人妻av无码中文专区|
亚洲国产综合精品2020|
人妻熟妇乱又伦精品视频|
国产亚洲精品久久久闺蜜|
欧美性生交A片免费看|
色综合色欲色综合色综合色乛|
无码免费一区二区三区|
亚洲欧美另类成人综合图片|
日韩人妻无码精品一专区二区三区|
精品无码人妻一区二区三区品|
国产人A片在线乱码视频|
国产乱沈阳女人高潮乱叫老|
最近中文字幕MV在线MV|
亚洲色欲色欲WWW在线看|
www.亚洲最大夜色伊人|
日韩AV一区二区精品不卡|
亚洲精品无码午夜福利理论片|
免费观看国产女人高潮视频|
又长又大又粗又硬3p免费视频|
香蕉久久人人97超碰caoproen|
中文无码伦av中文字幕|
国产精品186在线观看在线播放|
成人h动漫精品一区二区|
久草原精品资源视频|
在线看片无码永久免费视频|
久久99精品国产麻豆蜜芽|
久在线精品视频线观看|
樱花草视频www日本韩国|
最近2018免费中文字幕4|
欧美性受xxxx视频|
国产日产欧产美韩系列麻豆|
俺来也俺去啦久久综合网|
亚洲中文有码字幕日本第一页|
人人爽人人爽人人片Av|
а的天堂网最新版在线|
日韩精品无码综合福利网|
亚洲成国产人片在线观看|
中国熟女仑乱hd|
97久久婷婷五月综合色D啪蜜芽|
精品国产一区二区三区久久
|
国产强奷伦奷片|
亚洲制服丝袜av一区二区三区|
8090成人午夜精品无码|
国产精品国产三级国产AV剧情
|
亚洲国产精品无码久久久五月天|
亚洲综合熟女久久久40p|
不卡av中文字幕手机看|
免费无码专区毛片高潮喷水|
久久只精品99品免费久23|
制服 丝袜 人妻 专区一本|
亚洲 另类 日韩 制服 无码|
亚洲伊人久久大香线蕉综合图片|
日韩中文高清在线专区|
亚洲人午夜射精精品日韩|
99久久精品国产自在首页|
丁香婷婷六月综合交清|
久久99精品国产麻豆婷婷洗澡
|
久久大香伊蕉在人线免费|
日日摸日日碰人妻无码|
中文字幕无码肉感爆乳在线|
黄乱色伦短篇小说h|
国产女同疯狂作爱系列|
日韩精品视频一区二区三区|
浪潮色诱av久久久久久久|
最近在线更新8中文字幕免费|
一区二区三区午夜无码视频|
天堂va欧美va亚洲va好看va|
国产艳情熟女视频|
国产成人Av一区二区三区不卡|
日本一区二区三区不卡免费|
给我免费播放片高清在线观看中国|
久久人妻无码一区二区|
麻豆国产人妻欲求不满|
亚洲一久久久久久久久|
日韩精品视频一区二区三区|
一 级 黄 色 片免费网站|
一二三四在线视频观看社区|
国产欧美成人xxx视频|
欧美性xxxxx极品少妇|
A4YY午夜福利视频无码|
婷婷综合久久中文字幕|
国产精品视频一区二区三区无码|
无码专区—VA亚洲V天堂麻豆|
日韩精品东京热无码视频|
乱人伦中文无码视频在线观看|
久久99久久99精品免视看|
久久无码国产专区精品|
日本激情在线看免费观看|
国产欧美va欧美va香蕉在线|
国产旡码高清一区二区三区|
亚洲区激情区无码区|
伊人久久大香线蕉av五月天|
精品一区二区三卡四卡网站|
精品无码日韩一区二区三区不卡|
国产人澡人澡澡澡人碰视频|
国内精品玖玖玖玖电影院|
国产又黄又爽又刺激的免费网址|
中日文字字幕乱码视频|
欧美人与zoxxxx视频|
成人性生交大免费看|
亚洲熟妇少妇任你躁在线观看无码|
国产亚洲欧洲997久久综合|
无套内谢少妇毛片A片流出白浆|
久久男人av资源网站无码|
东京热无码人妻一区二区AV|
色婷婷小说|
精品一区二区三区在线成人|
成本人妻片无码中文字幕免费|
人妻用嘴含精大口吞精|
久久综合给合久久国产免费
|
国语熟妇乱人乱A片久久|
中国老肥熟女BBW视频|
国精产品一区一区三区免费视频|
久久久久久国产精品免费无码|
国产精品久久久亚洲|
熟妇女人妻丰满少妇中文字幕|
色欲av无码一区二区三区|
欧美性猛交XXXX|
免费无限次永久看黄的APP|
在线播放国产女同闺蜜|
亚洲午夜久久久久久久久久|
亚洲日韩日本中文在线|
成人无码潮喷在线观看|
无码8090精品久久一区|
a篇片在线观看网站|
午夜久久久久久禁播电影|
玩弄白嫩少妇xxxxx性|
精品一卡2卡三卡4卡免费网站|
啦啦啦中文在线视频免费观看|
juliaann精品艳妇|
久久亚洲国产精品五月天婷|
成人无码视频在线观看网站|
精品一卡2卡三卡4卡免费网站|
999ZYZ玖玖资源站永久|
亚洲人成色99999在线观看|
无码137片内射在线影院|
激情内射人妻1区2区3区|
A片扒开双腿猛进入免费观看|
久久国产精品无码网站|
好男人在线社区www|
中国极品少妇XXXXX|
疯狂做受XXXX国产|
亚洲国产精彩中文乱码av|
人妻系列无码专区久久五月天|
宅宅少妇无码|
亚洲在AV极品无码|
人妻少妇无码精品视频区|
欧美FREESEX8一10精品|
青草青草久热精品视频在线观看
|
精品国产青草久久久久福利|
久久99精品国产麻豆宅宅|
人人妻人人妻人人人人妻|
国产精品麻豆欧美日韩ww|
性久久久久久|
国产制服丝袜亚洲日本|
伊人久久大香线焦av色|
亚洲国产区男人本色在线观看|
久久精品国产中国久久|
陈少霞三级|
精品国产yw在线观看|
久久99精品国产麻豆蜜芽|
精品久久久久久久免费人妻|
爽爽影院免费观看视频|
欧美精品一卡2卡3卡4卡乱码|
欧美牲交A欧美牲交aⅴ|
国产在线无码视频一区|
久久亚洲国产精品五月天婷|
国产成人久久婷婷精品流白浆|
狠狠色狠狠色综合网老熟女|
国产交换配乱婬视频A|
国产午夜成人免费看片|
国色精品无码专区在线不卡|
亚洲精品无码久久久久去q|
国产福利一区二区三区高清|
内射合集对白在线|
777久久精品一区二区三区无码|
少妇性BBB搡BBB爽爽爽|
97人妻超在线观看免费|
亚洲αv久久久噜噜噜噜噜|
亚洲中文字幕伊人久久无码|
成人欧美一区二区三区1314|
无码人妻丰满熟妇区96|
中国女人内谢69XXXX免费视频|
国产精品视频熟女韵味|
亚洲一区无码精品色|
丰满人妻熟妇乱又伦精品|
国产乱子影视频上线免费观看|
亚洲成av人片在www鸭子|
国产激情无码一区二区三区|
最近中文2019字幕第二页|
日韩精品一区二区三区色欲AV|
亚洲国产综合另类视频在线观看|
国产精品无码无卡无需播放器|
免费看欧美成人A片无码|
亚洲ⅴ国产v天堂a无码二区|
大地影视资源官网第二页|
亚洲国产精品18久久久久久|
三年成全免费高清大全|
99精品国产在热久久婷婷|
漂亮少妇高潮A片XXXX|
色综合天天综合网中文|
香蕉成人伊视频在线观看|
纯肉无遮挡日本动漫视频在线观看|
国产成A人亚洲精品无码樱花|
国产精品毛片在线完整版sab|
夫では満足できない人妻|
美女视频黄是免费|
性无码一区二区三区在线观看|
成年女人免费毛片视频永久vip
|
精精国产xxxx视频在线|
国模gogo无码人体啪啪|
亚洲人成网亚洲欧洲无码|
污草莓樱桃丝瓜绿巨人秋葵ios|
丰满少妇高潮惨叫久久久|
亚洲VA久久久噜噜噜久久|
国产超碰人人做人人爽AV|
国产黄大片在线观看画质优化
|
影音先锋人妻av在线电影|
热re99久久精品国99热|
已满18点此自动转m329|
国产免费观看久久黄AV片|
色综合视频一区中文字幕|
妇女敕bbb搡bbbbbb搡|
激情综合婷婷丁香五月尤物|
国产极品JK白丝喷白浆羞羞|
好男人视频社区2018年|
国产欧美久久久精品影院|
男JI大巴进入女人的视频小说|
中文字幕乱码无码人妻系列蜜桃|
少妇做爰喷水高潮呻吟A片免费|
亚洲精品国产第一区第二区|
欧美成人www免费全部网站|
欧美日韩精品|
337p日本大胆欧洲色噜噜|
国产精品民宅偷窥盗摄|
99精品国产兔费观看久久99|
亚洲在线|
亚洲色成人网站WWW永久男男|
欧美牲交A欧美牲交VDO|
菠萝蜜视频在线观看入口|
日韩AV无码成人网站|
最近2018中文字幕免费看2019|
久久综合av色老头免费观看|
亚洲国产高清国产精品|
最近免费中文字幕大全免费版视频|
日韩精品一区二区在线观看|
草蜢网在线观看免费高清|
免费人成自慰网站|
国内精品视频一区二区三区|
8488tv直播nba免费观看|
久久在精品线影院|
日本乱偷人妻中文字幕在线|
亚洲国产成人精品无码一区二区三区|
亚洲乱码中文字幕综合|
无码丰满熟妇|
国产69精品久久久久APP下载|
成全影院电视剧在线观看|
麻豆精品导航|
97久久精品人人槡人妻人人玩|
丰满人妻连续中出中文字幕在线|
色婷婷五月综合亚洲小说|
亚洲欧美日韩高清一区|
丝袜美腿亚洲一区二区|
国产乱子轮XXX农村|
成人a大片在线观看|
18款禁用软件app糖心vlog|
蜜臀AV无码国产精品色午夜麻豆
|
最近免费2019中文字幕大全
|
永久在线观看免费视频|
国产旡码高清一区二区三区|
一本大道精品视频在线|
亚洲欧美一区二区三区日产|
yy亚洲人成电影网站色|
欧美人和黑人牲交网站上线|
国产女人18毛片水真多|
中文天堂最新版在线|
Japanese极品少妇|
国内精品久久久久久99蜜桃|
深爱五月综合缴情综合网|
久久精品国产亚洲a∨麻豆|
日本久久综合久久鬼色|
国产亚洲精品美女久久久久|
日韩亚洲国产中文字幕欧美|
综合无码一区二区三区四区五区|
亚洲欧美日韩高清一区|
午夜色大片在线观看免费|
五级黄高潮片90分钟视频|
性欧美牲交xxxxx视频αpp|
亚洲精品国产二区图片欧美|
国精品无码一区二区三区在线|
中文人妻无码一区二区三区|
日韩人妻无码一区二区三区综合|
亚洲另类无码专区丝袜|
国产AV无码专区亚洲AⅤ|
欧洲无码八A片人妻少妇|
高清亚洲日韩欧洲不卡在线|
狠狠躁日日躁夜夜躁A片|
免费高清自慰区|
玩弄丰满少妇高潮A片推油小说|
少妇偷倩打野战|
在线看免费无码的av天堂|
亚洲国产制服丝袜无码av|
吸咬奶头狂揉60分钟视频|
国产亚洲精选美女久久久久|
亚洲综合天堂婷婷五月
|
日本无码不遮挡又黄大尺度电影|
爽爽精品dvd蜜桃成熟时电影院|
中文字幕精品AV一区二区五区
|
久久精品中文字幕大胸|
国精产品一区一区三区免费视频
|
日本三级在緌观看|
久久艳务乳肉豪妇荡乳A片|
99RE热视频这里只精品|
狠狠色噜噜狠狠狠8888米奇|
99精品全国免费观看视频
|
乱子伦av无码中文字幕|
特级A欧美做爰AAAAA片|
欧美亚韩一区二区三区|
97久久香蕉国产线看观看|
体检强奷小箩莉小说|
国产白嫩漂亮美女在线观看|
中文字幕亚洲情99在线|
亚洲日韩中文字幕在线播放|
少妇高潮呻吟A片免费看小说
|
午夜影院app在线1|
国产亚洲情侣一区二区无
|
色久悠悠婷婷综合在线亚洲|
欧美激情乱人伦|
日本护士毛茸茸高潮|
中国老肥熟女BBW视频|
亚洲欧洲美洲无码精品Va|
人妻丰满熟妇岳av无码区HD|
女人高潮特级毛片|
无码熟妇人妻AV在线影片最多|
精品国产制服丝袜高跟|
婷婷五月深深久久精品|
1000部精品久久久久久久久
|
色欲人妻AAAAAAA无码|
国产色精品久久人妻无码看片|
青青青伊人色综合久久|
中文字幕丰满伦子无码|
日韩一区二区三区无码影院|
亚洲欧美精品伊人久久|
国产精品国产自线拍免费|
国产又A又黄又潮娇喘视频|
欧美精品一国产成人综合久久|
2020亚洲国产精品无码
|
亚洲国产三级在线观看|
日本欧美一区二区三区高清|
少妇人妻偷人激情视频|
国产三级精品三级在线专1|
激情欧美日韩一区二区|
性大毛片视频|
亚洲精品国产综合久久久久紧|
免费欧洲美女牲交视频|
欧美交换配乱吟粗大25p|
精品aⅴ老司机天堂网站|
久久午夜福利电影|
久久精品国产久精国产69
|
欧美亚洲国产成人一区二区三区|
日本55丰满熟妇厨房伦|
手机无码人妻一区二区三区免费|
97精品伊人久久大香线蕉APP|
国产精品WWW夜色视频|
亚洲日韩欧洲无码a∨夜夜|
亚洲日韩A∨无码久|
五月激激激综合网亚洲|
麻豆人人妻人人妻人人片av|
伦人伦XXXX国语对白|
亚洲一区二区三区在线观看网站|
五月综合网亚洲乱妇久久|
日韩精品久久无码人妻中文字幕|
上司人妻互换HD无码中文|
精品久久久久久无码国产|
亚洲熟妇丰满大屁股熟妇
|
日韩精品久久无码中文字幕|
阿阿痛痛痛痛痛痛痛痛痛疼疼疼疼|
男人猛躁进女人的毛片A片|
久久国产精品人妻丝袜|
国产精品沙发午睡系列999|
国产在线视欧美亚综合|
精品国产成人一区二区|
99精品人妻少妇一区二区|
借妻中文字幕高清|
最好看高清免费视频|
伊人色综合一区二区三区|
亚洲欧美一区二区三区在线|
99国产精品成人片|
精品AV国产一区二区三区四区
|
免费无码又爽又刺激高潮软件|
日韩精品亚洲人成在线观看|
性色蜜桃臀x88av|
成人无码一区二区三区网站|
人妻av无码av中文av日韩av|
国产精品高清视亚洲中文
|
18禁男女无遮挡羞羞视频免|
大地资源二中文在线观看高清|
在厨房被c到高潮a毛片奶水|
国产精品夜夜春夜夜爽久久小
|
国内精品视频一区二区三区|
国产精品久久欧美一区|
久久国产一区二区三区|
亚洲人成网77777香蕉|
麻豆成人精品国产免费|
日韩精品久久久久久免费|
久草v在线视频免费观|
国产成人亚洲综合网站小说|
巨臀精品无码AV在线播放|
亚洲精品自在在线观看|
波多野结衣BT|
亚洲 日本 欧美 中文字幕|
最近最新高清中文字幕mv在线|
中文无码乱人伦中文视频在线v|
99久久国产综合精品五月天喷水|
精品乱人伦一区二区三区|
国产精品久久久久久久久久久久午衣片|
东北少妇不戴套对白第一次|
日韩Aⅴ人妻无码一区二区|
亚洲精品专区在线观看|
国产成人综合久久精品|
亚洲色欲色欲WWW在线看|
一线高清中文免费观看视频
|
最好看的2018中文2019|
日韩少妇内射免费播放|
亚洲国产精彩中文乱码av|
亚洲最大成人网色|
国产一区二区三区无码免费|
色五月丁香五月综合五月4438|
亚洲 欧美 动漫 少妇 自拍|
免费无码又爽又刺激高潮视频|
亚洲成av人片在线播放无码漫画|
久久男人Av资源网站无码软件|
给我免费播放片在线观看
|
99ER国产这里只有精品视频免费|
少妇人妻无码专区视频|
亚洲欧美精品AAAAAA片|
中文字幕亚洲一区二区va在线|
天堂岛最新版在线bt天堂|
亚洲一区二区三区无码中文字幕|
亚洲国产精品成人网址天堂|
亚洲 日韩 激情 无码 中出
|
午夜人妻久久久久久久久|
日本VA欧美VA精品发布|
√最新版天堂资源网在线|
免费观看欧美日韩亚洲|
东北少妇不戴套对白第一次|
国产欧美日韩一区二区三区|
少妇性做爰XXXⅩ性视频|
荡女精品导航|
伊人综合网站|
亚洲线精品一区二区三区影音先锋|
99精品国产在热久久无码|
亚洲国产精品+制服丝袜|
精品久久久久久久久久久aⅤ|
亚洲免费精品aⅴ国产|
久久久久亚洲国产AV麻豆|
日韩高清在线亚洲专区小说|
久久精品国产一区二区无码|
中国凸偷窥XXXX自由视频|
婷婷六月激情综合一区|
av在线无码专区一区|
国产精品久久久久久久久久妞妞|
亚洲精品一区二区三区福利|
婷婷四房综合激情五月在线|
国产福利一区二区三区在线视频|
中日韩亚洲人成无码网站|
欧美综合自拍亚洲综合图|
亚洲最大AV无码网址|
99国产精品国产精品九九|
国产精品麻豆欧美日韩ww|
97在线国内在线观看视频|
国产国拍亚洲精品AA片在线看|
《艳香艳史》在线播放|
免费的日本中文字幕视频|
中文字幕无线码一区二区|
中文字幕亚洲无线码在线一区|
国产色综合天天综合网|
中文人妻无码一区二区三区信息|
少女配狗视频大全播放|
国产网曝在线观看视频|
久久精品人妻无码专区|
精品久久久久久无码人妻蜜桃|
亚洲va久久久噜噜噜久久狠狠|
国自产拍在线网站|
eeuss鲁丝片AV无码|
华人在线亚洲欧美精品|
吃奶呻吟打开双腿做受视频|
久久综合久久久久88|
中文字幕熟女人妻佐佐木|
亚洲精品又粗又大又爽A片|
国产美女被遭强高潮露开双腿
|
女的被弄到高潮娇喘喷水视频|
久久国产主播福利在线|
羞国产在线拍揄自揄视频|
人人妻人人做人人爽夜欢视频
|
无码av中文一区二区三区桃花岛|
亚洲成av人片在线观看|
亚洲成av人片天堂网|
国产午夜人做人免费视频|
涩涩片大全百度影音|
久久久无码精品亚洲日韩按摩|
无码人妻21P|
国产精品永久久久久久久久久|
亚洲精品白浆高清久久久久久|
亚洲精品天堂无码中文字幕|
麻豆画精品传媒2021网站|
久久亚洲精品无码gv|
已满18点此自动转m329|
精品少妇人妻AV一区二区三区|
中文字幕乱码亚洲∧v日本|
亚洲情综合五月天|
欧美精品一区二区在线观看播放|
肉妇春潮干柴烈火MYFDUCC|
欧美伊人久久大香线蕉综合
|
亚洲中文字幕成人野花高清樱花|
性一交一乱一A片WWW|
亚洲国产福利成人一区二区|
亚洲精品无码高潮喷水在线|
久久精品a亚洲国产v高清不卡
|
(无码视频)在线观看|
欧美性受xxxx狂喷水|
夜夜躁日日躁狠狠久久av|
2019年国产精品手机视频|
18款禁用黄台视频软件下载入口|
黑人大战亚洲人精品一区|
少妇高潮A片无套内谢麻豆传|
国产精品网红主播美女|
成品网站w灬源码16|
久久综合网丁香五月|
日本在线视频WWW色影响|
日本欧美一区二区三区高清|
久久久国产精品人人片|
国产AV毛片亚洲含羞草社|
国产精自产拍久久久久久蜜
|
50岁熟妇高潮惨叫|
丰满女老板BD高清A片|
性饥渴艳妇性色生活片在线播放|
久久亚洲日韩看片无码|
超碰caopro熟女m超碰分类|
成年无码AV片在线无缓冲|
亚洲乱色伦图片区小说|
亚洲AⅤ在线无码天堂777|
亚洲 日韩 激情 无码 中出
|
色婷婷美国农夫综合激情亚洲
|
最近中文字幕高清中文字幕2018|
漂亮人妻被黑人久久精品|
精品无码久久久久国产|
国产精品538一区二区在线|
国产中文字幕乱人伦在线观看|
亚洲人成在线观看网站无码|
又色又爽又黄的A片免费看苍井空
无码日本邻居大乳人妻波多野结衣
|
欧美精品中文字幕在线视|
最近中文字幕MV高清在线|
人妻少妇中文字幕久久|
亚洲日韩在线中文字幕综合|
免费人成在线观看视频播放|
亚洲va天堂va欧美ⅴ|
中文字幕人妻被公上司喝醉|
国产成人精品久久综合|
亚洲成av人片无码天堂下载
|
亚洲综合色在线视频WWW|
丁香婷婷色综合激情五月
|
东京热无码人妻一区二区AV|
老太性开放bbwbbwbbw|
日韩人妻一区二区三区免费|
人妻丰满熟妇av无码区app|
重囗味sm在线观看无码视频|
中文字幕乱码无码人妻系列蜜桃|
色爱无码A V 综合区|
国内精品自在自线视频|
99草草国产熟女视频在线|
国产香蕉尹人在线视频你懂的|
国产精品无码无片在线观看3D|
丁香花在线视频观看免费|
а√天堂资源中文在线官网|
亚洲日韩成人AV无码网站
|
免费人成视频x8x8入口AA|
久久综合给合久久狠狠狠974
|
少妇人妻真实偷人精品视频|
亚洲成AV人片看电影AV|
琪琪女色窝窝777777|
freesex性中国熟妇|
9.11免费网站nba|
永久免费的无码中文字幕|
国产精品兄妹在线观看麻豆|
国产片a国产片免费看视频|
男人添女人荫蒂视频观看|
无人区AV在线观看|
精品国产欧美一区二区|
99久久亚洲综合精品成人网|
亚洲精品久久国产高清情趣|
www.色五月|
无码人妻精品一区二区三区9厂|
亚洲精品国产精品乱码视色|
国内高清在线观看视频|
97色碰碰公开视频|
免费香蕉成视频人网站|
狠狠色狠狠色五月激情|
午夜精品一区二区三区在线观看|
人妻有码中文字幕|
免费无码又爽又刺激高潮的视频|
男插女高潮一区二区|
亚洲欧美在线x视频|
国产一区二区三区在线观看免费
|
无码专区中文字幕无码|
曰本无码人妻丰满熟妇5g影院
|
亚洲国产精品成人无码区|
性VIDEOSGRATIS灌满|
国精品无码一区二区三区左线|
国产高清视频在线观看69|
国产亚洲日韩在线三区|
班花在教室轮流澡到高潮视频|
亚洲人护士毛茸茸|
国产av夜夜欢一区二区三区|
亚洲第一AV网站|
五月丁香六月狠狠爱综合|
狠狠躁夜夜躁青青草原软件|
亚洲高清国产拍精品26u|
99久热国产精品视频尤物|
国产精品一国产AV麻豆|
亚洲欧美日韩国产精品一区|
久久综合九色综合国产|
国内少妇毛片视频|
久草日b视频一二三区|
日韩无码专区|
亚洲αv久久久噜噜噜噜噜|
99热这里只有精品免费播放|
免费XXXX大片国产片|
激情五月亚洲综合图区|
成人免费无码大片a毛片|
亚洲欧洲自拍拍偷精品网314|
影音先锋男人AV橹橹色|
无码精品少妇在线人妻AV|
色窝窝无码一区二区三区成人网站|
国产午睡沙发系列|
久久人人爽人人爽人人av|
性欧美牲交xxxxx视频αpp|
亚洲精品国产二区图片欧美|
国产一区二区波多野结衣|
最新版天堂中文在线|
亚洲精品嫩草研究院久久|
国产美女亚洲精品久久久综合|
久久国产亚洲精品超碰热|
天天做天天大爽天天爱|
国产成人综合久久精品推荐|
国产片a国产片免费看视频|
丁香五月亚洲综合在线|
国产精品自在拍在线播放|
亚洲无人区电影国产|
亚洲另类春色国产精品|
四虎永久在线精品视频|
最近最新高清中文字幕mv在线|
亚洲色欧美色2019在线|
亚洲成av人片一区二区三区|
无码人妻少妇久久中文字幕|
qd11.878skins|
国产又粗又爽又猛的视频A片|
久久无码精品一区二区三区|
荡女精品导航|
中文字幕人妻熟在线影院|
人妻少妇不满足中文字幕|
久久视热在线视频精品|
国产在线观看免费视频在线|
日本熟妇色熟妇在线视频播放|
亚洲国产精品成人网址天堂|
国产成人亚洲精品无码AV大片|
狠狠躁夜夜躁人人爽天天5|
亚洲色无码综合图区手机|
国产成人精品视频a片免费网站|
4438丁香五月亚洲最大成人|
国产成人无码a区在线观看视频|
日韩无套内射视频6|
国产精品18禁污污网站|
无码国内精品久久综合88|
国产精品露脸国语对白|
精品国产乱码久久久软件下载|
狠狠色综合播放一区二区|
免费无码鲁丝片一区二区|
男女裸体做爰视频免费|
欧美国产精品日韩在线|
国产成人AV乱码免费观看|
无码国产69精品久久久久孕妇|
日本又黄又爽gif动态图|
国产精品亚洲日韩欧美色窝窝色欲|
精品国产乱码久久久久久浪潮|
人人妻人人澡人人爽国产|
苍井空亚洲精品AA片在线播放|
久久久久久亚洲精品|
欧美高清在线精品一区|
久久国产加勒比精品无码|
国产成人av免费观看|
国产精品一区二区在线观看99|
国产成人精品无码一区二区|
性盈盈网站久久久久忘忧草|
妇乱子伦毛片视频|
人妻无码一区二区三区免费|
成人网站色情WWW在线|
国色天香色欲色欲综合网|
午夜亚洲影院在线观看|
2022国产成人精品视频人|
卡一卡二卡三免费视频|
草蜢网在线观看免费高清|
国产午夜亚洲精品理论片不卡|
性无码专区无码|
国产国拍亚洲精品永久69|
JVID亚洲精品无圣光图套|
欧美人妖aa1片|
久久久久久夜精品精品免费啦|
人妻无码中文字幕一区二区三区|
国内精品国产成人国产三级|
狼色精品人妻在线视频免费|
中文字幕无码免费久久|
国产精品乱子乱xxxx|
日韩国产成人无码av毛片|
色丁狠狠桃花久久综合网
|
国产成人涩涩涩视频在线观看|
亚洲乱色熟女一区二区三区丝袜|
成人亚洲欧美一区二区|
丰满熟妇人妻av无码区|
年轻的妺妺2伦理HD中文字幕
|
久久亚洲精品无码Va白人极品|
中文字幕色AV一区二区三区
|
亚洲国产精品尤物YW在线观看
|
日产乱码芒果视频|
黄桃AV无码免费一区二区三区|
嫖妓丰满肥熟妇在线精品|
无码又爽又刺激A片涩涩动漫软件
少妇搡BBBB搡BBB搡澳门
|
男人和女人做爽爽视频|
中文最新版地址在线|
鲁一鲁亚洲无线码|
色偷偷人人澡人人爽人人模|
亚洲国产成人av毛片大全|
国产精品自在线拍国产|
中文字幕日韩人妻不卡一区|
国产男女猛烈无遮挡免费视频|
少妇人妻真实偷人精品视频|
亚洲国产精品福利片在线观看|
www插插插无码视频网站|
色欲av无码一区二区三区|
av色综合网站|
污污免费看锕锕锕锕锕锕|
国产成人无码A区精油按摩|
久久久久久欧美精品无码超碰|
国内精品久久人妻无码妲己影院|
国产亚洲成av人片在线观黄桃|
亚洲国产精品无码久久一线|
又爽又黄又无遮挡网站|
青青青草国产线观|
狠狠CAO2020高清视频|
国产最新免费高清在线视频|
丰满人妻一区二区三区无码Av
|
国产在线拍揄自揄视频网站|
国产偷v国产偷v亚洲高清|
欧美精品18videosex性欧美|
永久黄网站色视频免费|
99精品视频在线观看|
WWW免费视频在线观看播放|
成人亚洲欧美丁香在线观看|
久久精品高清一区二区三区|
99久久国产综合精品1|
国产AV无码熟妇人妻麻豆|
亚洲 自拍 另类 欧美 综合|
亚洲中文字幕琪琪在线|
麻花传媒MV一二三区别在哪里|
国产午夜福利在线播放87|
强奷乱码欧妇女中文字幕熟女|
免费人成年激情视频在线观看|
亚洲熟妇无码久久精品|
秋霞无码AV久久久精品|
久久人妻av无码中文专区|
人妻少妇伦在线无码专区视频|
少妇愉情理伦片丰满丰满午夜|
五月丁香综合激情六月久久|
四虎成人精品国产永久免费|
被义子侵犯的漂亮人妻中字
|
国产精品亚洲ΑV三区|
天堂а√在线最新版中文在线
|
国产精品亚洲а∨天堂免|
人人妻人人做人人爽|
最近中文字幕高清中文字幕2018
|
久久精品99久久久久久久久
|
色欲AV无码一区二区人妻|
欧美又硬又粗进去好爽A片
|
午夜福利在线永久视频|
四虎AV永久在线精品免费观看
|
国产午夜精品理论片|
中文字幕无码乱人伦免费|
影音先锋资源男人网|
妇乱子伦毛片视频|
亚洲一区二区三区无码中文A片
|
在线观看亚洲人成网站A片|
亚洲爆乳无码专区WWW|
久久精品国产清高在天天线|
中文字幕人妻av一区二区|
国产av成人一区二区三区|
伊人精品成人久久综合
|
国产精品一区二区国产主播|
18禁又污又黄又爽的网站不卡
|
女人自熨全过程直播|
chinese国产hd中国熟女|
48久久国产精品性色aⅴ人妻|
97久久精品无码一区二区天美|
国产午夜福利精品久久|
国产精品永久免费|
少妇内射视频播放舔大片
|
国产成人无码专区|
最近中文字幕电影免费mv|
久久无码欧美一二三区|
成人爱做日本视频免费|
日韩精品一区二区三区中文不卡
|
国产亚洲精品欧洲在线观看|
精品午夜福利1000在线观看|
国产一区二区三区日韩精品|
欧亚精品卡一卡二卡三|
99久久久无码国产精品性|
人妻有码中文字幕|
国内精品伊人久久久影视|
久久无码欧美一二三区|
粉嫩人国产呦系列(634)|
精品国产第一福利网站|
久久九色综合九色99伊人|
久久精品国产亚洲a∨麻豆|
国产成人一区二区三区|
日韩不卡手机视频在线观看|
无码人妻专区免费视频|
久久久久无码精品国产不卡|
麻婆豆腐传媒视频免费|
仓井空电影大全百度影音|
好男人在线社区www在线影院|
老鸭窝视频在线观看|
日韩精品人妻系列无码专区|
国产裸体美女永久免费无遮挡|
色偷偷尼玛图亚洲综合|
热99RE6久精品国产首页青柠|
国产免费午夜福利蜜芽无码|
少妇高潮A片无套内谢麻豆传|
9.1免费网站nba在线观看|
国产粉嫩高中无套进入|
国产精品视频第一区二区三区|
性色欲网站人妻丰满中文久久不卡|
国产亚洲精品久久无码98|
久久精品中文字幕无码绿巨人|
亚洲精品一区二区绿巨人|
性Free老太婆性XXX|
人妻av乱片av出轨av|
丰满熟妇乱子伦|
五月香丁激情欧美啪啪|
国内少妇毛片视频|
67194免费视一频观看|
国产av巨作丝袜秘书|
精品一卡2卡三卡4卡免费网站|
99riav国产精品视频|
国产福利一区二区三区在线观看
|
18禁无遮挡无码网站免费|
少妇熟女久久综合网色欲|
久久精品无码午夜福利理论片|
少妇无码一区二区三区|
中文成人无码精品久久久|
www插插插无码视频网站|
97精品国产手机|
秋霞AV一区二区二三区|
99久久国产综合精品麻豆|
久久国产精品二国产精品|
亚洲熟妇AV一区二区三区下载|
国产一区二区三区免费播放|
日韩亚洲欧美中文在线|
香蕉久久AV一区二区三区|
日本红怡院一本道|
荡公乱妇HD在线播放BD|
高潮VPSWINDOWS国产乱|
国产成人精品A视频|
精品一区二区不卡无码av|
把腿张开老子臊烂你多P视频
|
97香蕉超级碰碰碰久久兔费|
国产成年女人特黄特色毛片免
|
亚洲精品成人av在线|
GOGOGO手机高清在线观看|
久久综合久久鬼色|
国产精品久久久久久久免费看|
国产精品自在拍在线播放|
AV在线网站无码不卡的|
国产精品卡一卡2卡三卡网站|
亚洲熟妇中文字幕五十中出|
99久久国产综合精品五月天喷水|
亚洲成A人V欧美综合天堂麻豆|
17·c18起草视频|
欧美色就是色|
丰满大爆乳波霸奶|
亚洲国产精品久久久久婷婷软件
|
99国产精品人妻无码网站|
亚洲中文久久精品无码ww16|
国产午夜精品一区二区|
秋霞鲁丝片av无码|
激情射精爆插热吻无码视频|
97夜夜澡人人爽人人模人人喊|
亚洲一区二区三区中文字幕在线|
亚洲成av人片不卡无码久久|
荡公乱妇HD在线播放BD|
欧美伊人久久大香线蕉综合|
√最新版天堂资源网在线|
内射一区二区精品视频在线观看|
丰满人妻连续中出中文字幕在线|
少妇2做爰A片|
日产精品久久久一区二区
|
无码人妻一区二区三区线|
久久久久久久久毛片精品|
欧美激欧美啪啪片sm|
色窝窝无码一区二区三区色欲|
秋霞国产成人精品午夜视频APP|
国自产拍在线网站|
久久精品无码一区二区无码|
gogogo高清视频大全|
性色av无码无在线观看|
中文幕无线码中文字蜜桃|
极品熟妇大蝴蝶20P|
Y111111国产精品久久久|
精品久久久久久无码专区不卡|
亚洲色欲AV无码成人专区|
国产+爽+黄|
亚洲人毛茸茸bbxx|
国产亚洲精品久久无码98
|
亚洲中文字幕无码永久在线不卡|
直接看的成人无码视频网站|
亚洲国产制服丝袜无码av
|
玩弄白嫩少妇xxxxx性|
国产精品无码午夜免费影院
|
狠狠色综合播放一区二区|
亚洲成在人线在线播放无码|
伊人情人色综合网站|
国产小视频在线观看网站|
粉嫩小泬无遮挡久久久久久|
国产AV无码一区二区二三区J|
日韩精品亚洲人成在线观看|
亚洲熟妇av日韩熟妇av|
少妇被又大又粗又爽毛片|
久久人妻无码一区二区|
欧美老熟妇乱xxxxx|
亚洲中文字幕无码爆乳|
国产无遮挡裸体免费直播|
性xxxx视频播放|
亚洲人成亚洲人成在线观看
|
无码日韩做暖暖大全免费不卡
|
国产av巨作丝袜秘书|
国产又色又爽又黄又免费的小说|
主人给我戴上奶牛榨乳器调教|
337p日本大胆欧洲色噜噜|
狠狠躁日日躁夜夜躁A片
|
欧洲AV无码放荡人妇网站|
国产精品美女www爽爽爽视频|
国产av巨作丝袜秘书|
97SE狠狠狠狠狼鲁亚洲综合色|
中文精品无码中文字幕无码专区|
少妇高潮A片无套内谢麻豆传|
丰满少妇人妻久久久久久|
国产成人亚洲精品青草天美|
最近高清中文在线字幕在线观看|
狠狠色狠狠色综合|
久久精品高清一区二区三区|
亚洲精品无码久久毛片波多野吉衣
|
国产无遮挡又黄又爽又色|
亚洲日韩欧洲无码av夜夜摸|
中文字幕久久精品一二三区|
久久久久青草线蕉亚洲|
亚洲专区 AV 第一页 在线|
内射小寡妇无码|
精品久久久久久久无码人妻热|
中文字幕一精品亚洲无线一区|
精品无码人妻一区二区免费蜜桃
|
亚洲成a人片在线观看国产|
亚洲成A人片在线观看无码|
老汉与饥渴的寡妇bd|
丰满饥渴老女人hd|
精品国产第一福利网站|
午夜福利伦伦电影理论片在线观看|
中国精品少妇HD|
巨大欧美黑人XXXXBBBB|
久久精品99久久香蕉国产色戒|
日本道色综合久久影院|
国产鲁鲁视频在线观看
|
久久香综合精品久久伊人|
日韩精品视频一区二区三区|
精品系列无码一区二区三区
|
午夜做爰XXXⅩ性高湖视频美国|
欲色影视天天一区二区三区色香欲|
国产欧美日本亚洲精品一5区|
久久久精品久久久久久96|
亚洲色欲综合一区二区三区|
久久国产色AV免费观看|
亚洲狠狠婷婷综合久久|
亚洲人成亚洲精品|
黑巨茎大战欧美白妞|
色噜噜狠狠狠狠色综合久|
少妇与子乱在线观看|
又白又大的奶头A片免费网站视频|
四川丰满妇女毛片四川话|
精品日产一卡2卡三卡4卡自拍|
真实国产乱子伦对白视频|
人妻 自慰 偷窥 无码|
WWW成人国产高清内射|
少妇真实自偷自拍视频6|
中国女人做爰A片|
精品欧洲AV无码一区二区三区|
久久国产劲爆∧V内射|
日本精品久久无码影院|
99久久超碰中文字幕伊人|
精品国产AV 无码一区二区三区|
成人片免费视频在线播放|
一区二区三区高清AV专区|
2021在线精品自偷自拍无码|
无码人妻一区二区三区免费n鬼沢|
国产裸体歌舞一区二区|
亚洲a成人无码网站在线|
精品亚洲国产成人蜜臀av|
在线观看无码H片|
国产精品视频一区国模私拍|
国产精品无码制服丝袜|
亚洲 欧美 国产 日韩 精品|
色婷婷基地|
国产精品iGAO视频|
精品无码AV一区二区三区不卡|
亚洲熟妇无码一区二区三区|
国内揄拍国内精品少妇国语|
亚洲午夜久久久久久久久电影网|
日韩av片无码一区二区不卡电影|
欧美亚洲国产日韩一区二区|
日本熟妇japanese丰满|
国产免费无遮挡吸奶头视频|
国产深夜男女无套内射|
亚洲国产精彩中文乱码av|
2019天天吃天天香蕉|
免费岛国av动作片在线观看|
国产精品美女久久久浪潮av|
亚洲成a人片77777kkkkk|
嗯男人r啊唔h哈|
精品一区二区三区免费视频|
婷婷亚洲综合五月天小说|
亚洲国产A∨无码中文777|
精品无码国模私拍视频|
无码人妻视频一区二区三区|
国产成人精品人人做人人爽|
无码不卡av手机版免费|
婷婷激情综合色五月久久|
永久免费无码AV网站在线观看|
少妇BWBWBWBWBWBW|
一本久久a久久免费精品不卡|
东京热人妻系列无码专区|
亚洲第一区无码专区|
337p日本欧洲亚洲大胆色噜噜
|
久久青草精品38国产|
国产又色又爽又刺激在线播放|
国产国拍亚洲精品AA片在线看|
少妇厨房愉情理伦片视频|
久久久久人妻精品一区二区三区|
欧美人与禽2O2O性论交|
麻豆国产精品VA在线观看
|
国模无码视频一区二区三区|
国产丰满人妻一区二区三区|
亚洲蜜芽在线精品一区|
精品人妻无码一区二区三区|
亚洲欧美闷骚少妇影院|
成人亚洲精品久久久久软件|
狼群社区论坛在线观看|
最新的国产成人精品2021|
国产av永久无码青青草原|
国产成人精品三级在线影院|
欧美亚洲日本国产黑白配
|
无码国产精品一区二区色情男同
|
寂寞少妇做SPA按摩|
亚洲中文字幕无码久久2020|
亚洲中文字幕无码一区日日添|
国精产品无人区一码二码三蘑菇
|
波多野结av在线无码中文|
精品无人区一区二区三区|
国产精品一区二区AV不卡|
热re99久久精品国99热|
国产精品无码免费专区午夜小说|
免费看成人羞羞视频网站在线看
|
国产SUV精品一区二区6|
亚洲 自拍 另类小说综合图区|
国产人A片777777久久|
国产精品A久久久久久久久|
免费看饥渴难耐的少妇软件|
国产欧美亚洲精品第二区软件
|
免费国产黄网站在线观看|
精品av天堂毛片久久久|
欧美乱子伦XXXX在线观看|
婷婷伊人五月尤物|
精品国产欧美一区二区|
亚洲精品国产一区二区色欲影院
|
蜜芽AV无码精品国产午夜|
久久AV无码AⅤ高潮AV喷吹|
一进一出男女啪啪120秒试看
|
日本少妇无码精品12p|
国产voyeur精品偷窥222|
国产偷国产偷亚洲高清人乐享|
亚洲国产精品久久久久久|
亚洲产国偷v产偷v自拍色戒|
亚洲小说区图片区另类春色|
麻花豆精产三产区区别|
男女车车的车车网站W98免费|
久久亚洲国产成人精品性色
|
无码任你躁久久久久久老妇|
欧美乱妇高清无乱码在线观看|
又粗又大内射免费视频小说|
亚洲午夜久久久久久噜噜噜|
欧美精品亚洲精品日韩已满十八|
国产 成人 综合 亚洲 网站|
日韩精品人妻系列无码专区|
又黄又爽又色的视频|
国产一区二区三区免费播放|
久久国内精品自在自线400部|
日韩精品无码av成人观看|
无码国产精成人午夜视频不卡|
亚洲综合久久一区二区|
国产精品亚洲ΑV三区|
久久久国产精华液|
亚洲国产精品久久久久秋霞小|
大尺度性做爰激情视频王琳|
精品久久久无码中文字幕天天|
国产乱国产乱老熟300部视频|
国产亚洲欧美另类一区二区
|
日韩人妻无码精品一专区二区三区|
2020日本不卡一区二区视频|
WWW.美女激情电影.Com|
97人妻天天摸天天爽天天|
日韩精品一区二区av在线|
国产激情无码一区二区三区
|
狼色精品人妻在线视频免费|
精品AV国产一区二区三区四区
|
国产AV国片偷人妻麻豆|
国产高潮国产高潮久久久|
妺妺坐在我腿上勃起弄了视频|
国产午夜永久福利视频在线观看|
国自产精品手机在线观看视频|
暴躁少女10-16岁|
成人做爰视频WWW|
精品国产免费一区二区三区|
久久精品国产曰本波多野结衣
|
亚洲精品国产综合久久一线|
久久在精品线影院|
狠狠躁夜夜躁人人躁婷婷视频|
免费大黄网站在线观|
久久99精品久久久久久久不卡
|
亚洲午夜无码久久|
少妇高潮大叫好爽|
亚洲成色www久久网站|
亚洲 欧美 国产 制服 动漫
|
久久AV无码AⅤ高潮AV喷吹|
亚洲国产天堂久久综合226114
|
日本高清成本人视频一区|
国产人成精品香港三级在线
|
国产精品午夜小视频观看|
97人妻天天摸天天爽天天|
日日擼夜夜擼|
h国产小视频福利免费视频|
无人在线观看高清视频单曲直播|
久久久久人妻一区二区三区vr|
久久青草精品38国产|
日韩内射美女人妻一区二区三区
|
大地资源影院在线播放|
尤物精品国产第一福利网站
|
粉嫩虎白扒开小泬|
伊人久久亚洲综合影院首页
|
国产高清管线视频免费|
亚洲成AV人片看电影AV|
亚洲第一AV导航AV尤物|
中文无码乱人伦中文视频播放|
噼里啪啦免费看高清片|
日韩av无码久久精品免费|
√天堂中文www官网在线|
99在线在线视频观看|
欧美重口变态另类A片|
婷婷丁香五月中文字幕|
无码视频免费一区二区三区|
久久久精品人妻无码专区不卡|
亚洲精品婷婷无码成人a片在线|
国产婷婷一区二区三区|
亚洲精品无码你懂的|
国产美女裸体丝袜喷水视频|
水多多导航凹凸福利在线|
日韩精品内射视频免费观看|
BBWBBMBBWBBM少妇|
亚洲欭美日韩颜射在线二|
小sao货撅起屁股扒开c打视频
|
人人综合亚洲无线码另类|
亚洲精品电影院|
97人妻精品一区二区三区|
中文字幕人成无码免费视频|
日本黄页网站免费大全|
免费网站看GV片在线|
国产精品无码无卡在线播放|
久久精品亚洲日本波多野结衣|
好吊妞人成视频在线观看|
黑人无套内谢中国少妇|
日产亚洲一区二区三区|
日韩精品无码一区二区中文字幕|
国产真人免费无码AV在线观看|
亚洲国产精品13p|
久久精品国产亚洲无删除|
国产在线观看无遮挡无码AⅤ多人|
免费免费视频片在线观看|
少妇做爰免费视频网站|
玩弄放荡人妇系列AV在线网站|
亚洲成av人片一区二区梦乃|
白嫩少妇bbw撒尿视频|
凹凸精品视频国产品免费|
最近高清中文字幕免费mv视频7|
国产强被迫伦姧在线观看无码|
黑巨茎大战欧美白妞|
少妇做爰喷水高潮呻吟A片免费|
欧美激情在线播放|
重口老太大和小伙乱|
男女多P混交群体交乱A片小说|
吸奶头吸到高潮视频免费视频|
蜜桃av抽搐高潮一区二区|
久久九九有精品国产尤物|
久久精品无码专区免费东京热
|
未满十八勿入av网免费|
夜夜爱夜夜做夜夜爽|
亚洲国产综合精品2020|
妓院一钑片免看黄大片|
成人免费视频无码专区|
欧产日产国产精品精品|
92午夜福利757视频在线观看|
秋霞成人国产理论A片|
中国女人内谢69XXXX免费视频|
大地资源免费第二页|
日韩精品无码中文字幕电影|
国产草莓精品国产av片国产|
艳妇荡女欲乱A片在线观看|
无码人妻丰满熟妇啪啪|
精品久久久中文字幕人妻|
免费人成在线观看视频播放|
欧美精品日韩精品一卡|
强奷人妻日本中文字幕|
色翁荡息又大又硬又粗又爽
|
李宗瑞性侵照片全集|
伊人久久大香线蕉aⅴ色|
99精品久久精品一区二区|
日本乱偷人妻中文字幕|
久久精品无码专区免费|
妖精色av无码国产在线看|
精品人妻系列无码人妻不卡|
已满十八岁从此入戴耳机的意思|
黑人巨大白妞出浆|
国产亚洲精品俞拍视频|
九九久久精品国产|
狠狠综合久久AV一区二区|
又粗又硬又黄又爽的免费视频|
亚洲日韩精品国产一区二区三区|
厨房的春潮A片|
小草在线观看中文字幕|
精品深夜AV无码一区二区老年|
亚洲丰满熟女一区二区哦|
99热这里只有精品免费播放|
精品一区二区三人妻视频|
92午夜福利757视频在线观看|
人妻无码人妻有码中文字幕|
苍井空巨乳教师线观看|
国产乱国产乱老熟300部视频|
国产人久久人人人人爽|
欧美性猛交久久久乱大交|
AV激情亚洲男人的天堂国语|
曰本无码人妻丰满熟妇5g影院|
国产男生午夜福利免费网站|
800AV凹凸视频在线观看|
最近中文字幕在线中文视频|
夜夜爽一区二区三区精品|
欧美性做爰又大又粗又长|
大地资源在线观看免费官网|
日韩高清在线中文字带字幕|
人妻无码中文专区久久av|
久久久老熟女一区二区三区|
欧美成人视频|
亚洲深深色噜噜狠狠网站|
亚洲中文av一区二区三区|
精品国产污污免费网站|
久久精品日韩av无码|
丰满人妻一区二区三区无码Av
|
无码国产偷倩在线播放老年人|
国产精品自在线拍国产电影|
亚洲日本va午夜中文字幕一区|
久久国产乱子伦免费精品|
强插女教师av在线|
国产欧美久久一区二区三区|
天天躁日日躁很很躁2022|
国产亚洲精品无码不卡|
国产精品无码无片在线观看3D|
四虎永久在线高清国产精品|
性夜夜春夜夜爽AA片A|
国产亚洲精品久久久999无毒|
精品国产a∨无码一区二区三区|
国产激情无码激情A片软件
|
成人免费看www网址入口|
亚洲va中文字幕无码毛片|
人妻丰满熟妞AV无码区|
中国xxxxxl19dfm免费|
老牛精品亚洲成AV人片|
黑人大荫道BBWBBB高潮潮喷|
精品人妻av区乱码|
久久国产免费直播|
曰韩精品无码一区二区三区|
亚洲伊人久久综合成人|
成年视频xxxxx在线网|
亚洲熟妇丰满大屁股熟妇|
人妻少妇被猛烈进入中文字幕|
最好看的中文视频最好的中文|
午夜自产精品一区二区三区|
少妇无码av无码去区钱|
亚洲精品在看在线观看|
最近中文字幕高清2019中文字幕|
午夜少妇性影院私人影院在线|
99久久精品这里只有精品|
国产国拍亚洲精品AA片在线看
|
中国国语对白高潮A片|
青青热久免费精品视频在线播放|
亚洲综合色在线观看一区二区|
无码成人av在线一区二区|
亚洲乱码国产乱码精华|
女人高潮抽搐30分钟|
精品亚洲AⅤ无码一区二区三区|
亚洲成a人片在线观看www|
欧洲美女粗暴牲交免费观看|
精品无码久久久久久尤物|
久久香综合精品久久伊人|
医院人妻隔着帘子被中出|
乱中年女人伦AV三区|
狼友久久国产精品|
欧美欧洲成本大片免费|
草莓樱桃丝瓜绿巨人秋葵ios|
宅宅少妇无码|
亚洲午夜私人影院在线观看|
国产精品美女久久久免费|
上司人妻互换HD无码中文|
97色碰碰公开视频|
√天堂资源网最新版在线|
久久99ER热精品免费播|
美女极度色诱视频国产|
欧美怡红院免费全部视频|
国产欧美日韩一区二区加勒比|
成人伊人精品色XXXX视频
|
人人妻人人做人人爽夜欢视频|
国产精品久久二区二区|
亚洲情综合五月天|
在线看片免费人成视频播|
成人无码区在线观看|
97国产超碰一区二区三区|
国产呦系列呦交|
AV无码一区二区三区|
丰满少妇被猛烈高清播放|
伸进同桌奶罩里摸她胸作文|
久久婷婷成人综合色|
伊人久久大香线蕉aⅴ色|
艳妇乳肉豪妇荡乳av|
国产精品扒开腿做爽爽爽日本无码
|
美丽人妻被按摩中出中文字幕|
无码精品国产VA在线观看|
亚洲S久久久久一区二区|
亚洲精品成人片在线播放|
免费无码A片一区二三区|
内射小寡妇无码|
99精品国产在热久久无毒不卡
|
亚洲欧洲无码AV电影在线观看|
国产欧美在线一区二区三区
|
在线无码va中文字幕无码|
亚洲精品电影院|
理论片免费ā片在线观看|
无码中文字幕人妻在线一区|
香蕉久久人人97超碰caoproen|
成年午夜性视频|
国产亚洲精品久久无码98|
天堂…中文在线最新版在线|
另类内射国产在线|
综合在线视频精品专区|
国产成年无码av片在线|
琪琪女色窝窝777777|
亚洲综合无码无在线观看|
又色又爽又黄的A片免费看苍井空|
亚洲中文字幕久久精品无码VA
|
最近2018中文字幕大全免费|
内射老妇bbwx0c0ck|
国产aⅴ精品一区二区三区久久|
中文毛片无遮挡高潮免费|
强奷人妻日本中文字幕|
麻豆成人久久精品二区三区免费|
久久亚洲私人国产精品va|
三年在线观看免费大全|
国产精品无码专区av在线播放|
无码任你躁久久久久久老妇蜜桃|
亚洲精品国产品国语在线|
日本少妇A片又爽又黄|
日本激情在线看免费观看|
亚洲国产精品久久网午夜|
亚洲国产精品无码毛|
国产午夜亚洲精品午夜鲁丝片|
男女后式激烈动态图片|
琪琪午夜伦伦电影理论片A片
|
亚洲精品无码国产爽快A片|
久久综合久久鬼色|
国产av巨作情欲放纵|
中国极品少妇XXXXX|
国产又粗又硬又大爽黄老大爷视|
午夜福利无码免费专区|
边摸边脱吃奶边高潮视频免费|
人妻aⅴ无码一区二区三区|
亚洲色无码中文字幕在线|
小说区 图片区色 综合区|
成人毛片100免费观看|
我我色综合|
а√天堂资源8在线官网在线|
天堂中文资源在线最新版下载|
扌臿辶畐的姐弟从小|
亚洲国产精品无码中文LV|
国产成人精品日本亚洲|
女厕厕露P撒尿八个少妇|
a国产一区二区免费入口|
搡老熟女中国老太|
中文无码久久精品|
aV无码久久久久不卡网站蜜桃|
(无码视频)在线观看|
国产又色又爽又刺激视频|
四川丰满妇女毛片四川话|
国产女精品视频网站免费|
AV老司机亚洲精品天堂|
亚洲色欲色欲77777小说|
japanese厨房乱tub偷|
久久久橹橹橹久久久久高清|
性饥渴的欲女少妇|
人妻体内射精一区二区三区|
国精产品一区一区三区免费视频
|
丰满人妻连续中出中文字幕在线|
久久久噜噜噜久久中文福利|
人妻夜夜添夜夜无码AV|
精品无码一区二区三区在线
|
全肉高黄高h的小说推荐|
亚洲欧美日韩精品久久|
日本欧美一区二区三区高清|
一区二区人妻无码欧美|
一本一道精品欧美中文字幕|
国产精品人人妻人人爽|
国产精品无码AV不卡|
无遮挡又黄又刺激的视频|
精人妻无码一区二区三区|
久久老子午夜精品无码|
99亚洲精品卡2卡三卡4卡2卡|
国产A在亚洲线播放|
成人午夜视频一区二区无码|
国产欧美日本亚洲精品一5区|
人妻夜夜爽爽88888视频|
婷婷五月综合激情中文字幕
|
国产精品视频一区国模私拍|
《丰满的女邻居》电影|
大黑大巴大战欧洲美女图片|
日韩无套内射高潮|
中国chinese4p交换video|
亚洲精品无码一区二区色戒|
JAPANESE内射×××|
csgo暴躁妹妹高清|
天美传媒xxxxhd videos3|
熟女体下毛毛黑森林|
国产av无码专区亚汌a√|
国产在线视频精品视频|
变态拳头交视频一区二区|
8090成人午夜精品无码|
麻豆国产尤物av尤物在线看|
无码一区二区三区老色鬼|
宅女午夜福利免费视频在线观看|
17·c18起草视频|
99亚洲精品色情无码久久|
在线精品视频一区二区三四|
国产av永久无码青青草原|
国产最新AV在线播放不卡|
国产精品亚洲综合一区二区三区
|
亚洲乱妇老熟女爽到高潮的片
|
国产白丝JK捆绑束缚调教视频|
男女做爰的全部过程A片|
女人裸体让男人吃奶视频|
人人妻人人澡人人爽欧美二区
|
欧美精品18videosex性欧美|
欧美 偷窥 清纯 综合图区|
免费吃瓜爆料曝光独家揭秘|
国产欧美一区二区精品久久久|
日韩欧美中文字幕看片你懂的|
国产成人av电影在线观看第一页|
人人爽人人爽人人片a∨|
最新中文字幕av无码不卡|
激情综合五月丁香亚洲|
亚洲热妇无码AV在线播放|
免费人成再在线观看|
久久无码人妻国产一区二区|
日本一道人妻无码一区视频|
在线观看中文字幕不卡有码|
久久99这里只有是精品6|
日韩av一区二区三区免费看|
最近最新中文字幕大全免费版下载|
久久久久久久久无码精品亚洲日韩|
欧美日韩国产码综合二区|
精品一区二区av天堂|
国产精品爆乳在线播放第一人称|
久久精品国产精品亚洲蜜月|
小荡货哦灬啊灬用力啊快视频|
日韩人妻无码精品二专区|
欧美综合自拍亚洲综合图|
国产成人亚洲综合网色欲网久下载
|
国产乱人无码伦av在线a|
成人白浆超碰人人人人|
国产另类ts人妖一区二区|
少妇真实自偷自拍视频6|
AV成人午夜无码一区二区|
人妻人人澡人人添人人爽人人玩
|
7777精品伊人久久久大香线蕉|
在厨房抱住岳丰满大屁股韩国电影
|
国内少妇毛片视频|
欧美性狂猛xxxxx|
国产成人精品永久免费视频
|
亚洲色偷拍区另类无码专区|
婷婷五月深深久久精品|
丝袜老师你下面夹得我好爽视频|
久久丁香五月天综合网|
日韩 欧美 动漫 国产 制服|
国产成人精品亚洲日本在线观看|
欧美成人a在线网站|
国产高清在线精品一本大道|
国产乱码一区二区三区|
精品无码人妻被多人侵犯AV|
亚洲综合熟女久久久40p|
夜夜躁狠狠躁日日躁视频|
怮交小U女天堂HD|
女女女女女裸体处开bbb|
97亚洲色欲色欲综合网|
亚洲人成无码网www电影蜜芽|
亚洲精品无码国产爽快A片|
精品无人区一区二区三区|
擦老太bbb擦bbb擦bbb擦|
国产免费又色又爽又黄的视频软件|
欧美人与动牲猛交XXXXBBBB|
国产欧美久久一区二区三区|
中文字幕久久波多野结衣AV不卡|
国产视觉盛宴在线视频|
亚洲国产婷婷香蕉久久久久久|
精品国产男人的天堂久久|
国产精品国产自线拍免费软件|
天堂中文在线资源库用|
亚洲 精品 综合 精品 自拍|
国产美女a做受大片观看|
中文字幕热久久久久久久|
久久伊人蜜桃AV一区二区|
日本亚洲欧洲免费无码|
日韓無碼人妻不倫A片|
国产精品无码专区在线观看不卡|
久久伊人精品影院一本到综合|
免费国产VA在线观看|
欧洲-级毛片内射八十老太婆|
丝袜美腿一区二区三区|
免费 人妻 无码 不卡中文...|
偷偷做久久久久网站|
亚洲人成网站免费播放|
亚洲欧洲日产国产 最新|
影音先锋中文字幕人妻|
夜夜高潮天天爽欧美|
精品国产第一福利网站|
亚洲精品嫩草研究院久久|
国产成人精选视频在线观看|
久久国产亚洲精品无码|
丁香花免费完整高清观看|
亚洲日韩中文字幕一区|
开心久久婷婷综合中文字幕|
日韩精品无码一区二区三区不卡|
娇妻被老头杂交|
weyvv8国产的suv|
国产乱人伦AV在线麻豆A|
一线高清中文免费观看视频|
亚洲乱色熟女一区二区三区丝袜|
亚洲午夜福利精品无码不卡|
免费精品国产自产拍在线观看图片|
精品无码国产污污污免费网站国产|
人妻精品动漫h无码专区|
97夜夜澡人人爽人人模人人喊|
无码国产精品一区二区免费虚拟VR|
亚洲综合图色40p|
日本国产网曝视频在线观看|
国产自产在线视频一区|
国产一极内射視颍一|
国产精品精品自在线拍|
男生和女生一起很痛的轮滑鞋|
少妇真实自偷自拍视频6|
青青草视频免费观看|
亚洲精品无码久久久久去q|
免费国精产品自偷自偷免费看|
成人免费一区二区三区视频软件|
久久久久无码精品国产不卡|
最好看高清免费视频|
51国产黑色丝袜高跟鞋|
看全色黄大色大片免费久久|
国产强被迫伦姧在线观看无码|
亚洲色国产AV天堂|
国产成人久久婷婷精品流白浆|
国产白袜脚足J棉袜在线观看
|
97人伦影院A片在线观看
|
日韩夜夜高潮夜夜爽无码|
18禁黄网站免费|
在线无码免费的毛片视频|
国产成人综合亚洲看片|
亚洲 欧美 中文 日韩 综合
|
糖心vlog入口进入|
亚洲伊人久久综合成人|
国产99视频精品专区|
久久综合色另类小说|
国产精品丝袜亚洲熟女|
久本草在线中文字幕|
亚洲午夜福利精品无码不卡|
日本高清www午色夜在线视频|
久久九九久精品国产|
成人无码免费一区二区三区|
精品久久久久中文字幕APP|
台湾佬自拍偷区亚洲综合|
午夜成年男人免费网站|
日韩人妻高清精品专区|
精品福利一区二区三区免费视频|
97久章草在线视频播放|
老少配老妇老熟女中文普通话|
欧美人与动XXXXZ0OZ|
久久青青无码av亚洲黑人|
伊人久久大香线蕉avapp下载|
欧美猛交XX无码黑寡妇|
人妻少妇不满足中文字幕|
av片在线观看|
成熟人妻AV无码专区A片|
国产又色又爽又黄的免费|
99精品国产兔费观看久久99
|
鲁鲁鲁爽爽爽在线视频观看|
人妻无码一区二区三区免费|
亚洲精品国产电影|
欧美成人一区二区三区|
亚洲第一无码精品立川理惠|
99久久久精品免费观看国产|
又大又长粗又爽又黄少妇视频|
浪潮色诱av久久久久久久|
亚洲精品天堂无码中文字幕|
无套内谢少妇毛片A片AV
|
亚洲 日本 欧美 中文字幕|
精品无人区一区二区三区|
国产最新进精品视频|
亚洲熟妇AV一区二区三区下载|
无码人妻精品国产日韩电影|
婷婷五月综合国产激情|
97久久超碰成人精品网站|
99久久久无码国产精品性|
国产免费爽爽视频|
久久大香国产成人av|
国产午夜福利精品久久|
久久美利坚合众国AV无码|
国产成人免费高清激情视频|
国模大胆一区二区三区|
亚洲妇女水蜜桃AV网网站|
欲帝精品福利视频导航|
japanese国产在线观看|
国产精品亚洲а∨天堂免|
《艳香艳史》在线播放|
又大又爽又硬的曰皮视频|
国产午夜人做人免费视频|
亚洲精品无码乱码成人|
国产午夜手机精彩视频|
又大又爽又硬的曰皮视频|
美女极品粉嫩美鮑20p图|
97久久久亚洲综合久久88|
日本少妇按摩做爰2|
天堂AV无码大芭蕉伊人AV不卡|
精品人体无码一区二区三区|
精品久久8X国产免费观看|
夜蒲4三级做爰视频|
A片无码AV看免费大片在线喝奶|
亚洲精品专区在线观看|
久久国产色AV免费观看|
AV人摸人人人澡人人超碰妓女|
中文字幕亚洲乱码熟女一区二区|
伊人久久综合狼伊人久久
|
精品无码国模私拍视频|
在线天堂新版资源www在线|
久久久一本精品99久久精品88|
国产色婷婷亚洲999精品小说|
日韩精品无码一区二区三区免费
|
亚洲国产另类久久久精品黑人|
哈尔滨老熟女啪啪嗷嗷叫|
性一交一乱一美A片|
精品久久久久久国产潘金莲|
浓毛老太交欧美老妇热爱乱|
最新中文字幕av无码不卡|
真人做爰高潮视频免费|
欧美午夜精品一区二区三区电影|
中文字幕丰满乱孑伦无码专区|
九九精品无码专区免费|
亚洲日本一区二区三区在线|
色戒完整版视频哔哩哔哩|
а√天堂网www在线最新版地址
|
青春草在线视频免费观看|
99亚洲狠狠色综合久久位|
《色戒》未删减版电影在线|
a篇片在线观看|
欧美又大又粗无码视频|
中文在线А天堂|
凹凸18禁在线视频|
四库影院永久国产精品|
亚洲国产精品无码中文字满|
精品国产亚洲一区二区三区|
午夜福利国产成人a∨在线观看|
久久精品夜夜夜夜夜久久|
精品无码一区在线观看|
夜蒲4三级做爰视频|
香蕉久久一区二区不卡无毒影院|
免费观看一区二区三区|
忘忧草视频入口|
免费无码A片一区二三区|
亚洲色大成成人网站久久|
亚洲精品婷婷无码成人a片在线|
最近2019年日本中文免费字幕
|
伊人久久大香线蕉AV一区二区|
亚洲一区av在线观看无码叶玉卿
|
亚洲欧美综合精品久久成人网|
国产精品久久久亚洲|
波多野结衣VA无码中文字幕电影|
无遮挡粉嫩小泬久久久久久久|
老司机久久精品最新免费|
永久亚洲成a人片777777|
亚洲国产精品无码久久秋霞|
亚洲精品一、二、三区|
97人妻人人澡人人爽国产一|
天堂俺去俺来也WWW色官网|
美女裸体视频黄的免费|
久久精品手机观看|
野狼AV午夜福利在线|
avtt天堂网久久精品|
2020国产在线拍揄自揄视频
|
亚洲精品国偷拍自产在线观看蜜臀
|
伦埋琪琪深夜福利|
亚洲男男无套GV大学生|
最近高清中文在线字幕在线观看
|
中文无码av在线亚洲电影|
蜜桃人妻无码AV天堂三区|
国产成人综合久久精品推下载
|
中文字幕人妻色偷偷久久|
国产免费丝袜调教视频|
办公室玩弄娇喘秘书在线观看|
色偷偷一区二区无码视频|
成熟人妻换xxxx|
女人色毛片女人色毛片18|
免费无码Av一区二区三区|
日韩Aⅴ人妻无码一区二区|
亚洲美女又黄又爽在线观看|
制服丝袜中文丝袜AV|
蜜芽国产尤物av尤物在线看|
最近日本字幕免费高清|
亚洲中久无码永久在线观看同
|
久久国产免费福利永久|
99精品国产一区二区|
国内精品视频一区二区三区|
日韩精品久久久久久免费|
久久久影视文化传媒有限公司|
国产欧美久久久精品影院|
蜜臀av一区二区|
亚洲大尺度无码无码专线一区
|
天天摸天天做天天爽天天舒服|
国产一区二区三区无码免费|
欧美xxxxxbbbb|
精品人妻一区二区三区四区|
久久国产色AV免费看|
国产激情精品一区二区三区|
亚洲乱亚洲乱少妇无码99P|
天天爽夜夜爽人人爽|
人妻丰满精品一区二区A片|
又硬又粗又大一区二区三区视频|
当着全班面被C到高潮哭视频|
亚洲精品无码国产|
狠狠躁夜夜躁人人躁婷婷视频|
黑人无套内谢中国少妇|
阿娇艳z门照片无码av4i|
无码人妻丰满熟妇区五十路|
自在自线亚洲А∨天堂在线|
久久99精品久久久久久齐齐百度|
中文字幕久久久人妻无码|
国内精品一区二区三区在线观看|
大学情侣宿舍里啪视频|
日韩在线视频线观看一区|
18禁影库永久免费|
精精国产xxxx视频在线|
嫩草研究院久久久精品|
亚洲午夜精品一区二区
|
亚洲伊人久久大香线蕉|
高清一卡二卡三卡四免费|
8090成人午夜精品无码|
国产成人欧美视频在线观看|
久久国产精品免费一区二区三区|
窝窝人体色www|
国产精品久久久久久tv|
亚洲综合激情另类小说区|
yy8男人的天堂|
欧美极度丰满熟妇HD|
日本婬乱A片AAA毛片麻豆软件
|
久久国产精品无码网站|
国产成人综合久久精品|
人妻无码少妇一区二区|
伊人情人色综合网站|
国产精品麻豆VA在线播放|
后入内射欧美99二区视频|
国产极品久久久久极品|
无码精品视频一区二区三区|
精品欧美H无遮挡在线看中文|
丁香花视频在线观看|
精品久久久久久无码中文野结衣
|
日本少妇xxx做受|
东京热一精品无码av|
日韩无码专区|
亚欧乱色熟女一区二区三区|
色综合久久无码中文字幕app|
丁香花在线高清视频完整版观看|
日本乱子人伦在线视频|
亚洲性日韩精品一区二区三区|
女人与拘做受AAAAA片|
试看120秒做受小视频免费|
三级国产三级在线|
亚洲欧美日韩综合一区
|
人与性口牲恔配视频免费观看|
中文字幕日本人妻久久久免费|
日本亲与子乱人妻hd|
重口老太大和小伙乱|
最近2018中文字幕大全免费|
a在线视频|
中文天堂最新版在线|
60分钟刺激战场视频|
水蜜桃一三四专区|
人妻无码免费一区二区三区|
一本道高清在一线视频|
亚洲国产福利成人一区二区|
日本韩国三级aⅴ在线观看|
无码人妻丰满熟妇啪啪|
日韩精品无码av中文无码版|
亚洲人成电影网站 久久影视|
中文字幕人妻偷伦在线视频
|
亚洲中文字幕久爱亚洲伊人|
国产极品粉嫩馒头一线天AV|
激情综合一区二区迷情校园|
边亲边脱边捏胸视频|
国产强被迫伦姧在线观看无码|
无码一区二区三区免费|
亚洲国产精品无码中文字|
亚洲狠狠婷婷综合久久久久图片
|
护士脱了内裤让我爽了一夜视频|
国产偷人爽久久久久久老妇APP|
久久国产色AV免费观看|
国产青草视频在线观看|
国产精品视频露脸|
性无码专区无码片|
少妇熟女天堂网av|
99久久精品免费看国产|
无码少妇精品一区二区免费动态|
一本一道久久综合狠狠老|
Chinese老女人老熟妇HD|
强睡邻居人妻中文字幕|
欧美亚洲日本国产综合在线|
国产V综合V亚洲欧美久久|
成人精品一区二区三区中文字幕
|
精品少妇人妻AV免费久久洗澡|
国产欧美久久久精品影院|
永久免费无码日韩视频|
亚洲熟女少妇一区二区|
久久无码精品一区二区三区|
国产成人av|
在线A亚洲视频播放在线观看|
久久精品国产亚洲夜色av网站|
交换邻居波多野结衣中文字幕|
中文字幕AV高清片|
伊人久久久大香线蕉综合直播|
国产精品民宅偷窥盗摄|
国产成人精品永久免费视频|
玩爽少妇人妻系列|
已满十八岁免费观看电视剧最新章节|
成人无码WWW免费视频|
亚洲A∨国产AV综合AV网站|
久久人妻av无码中文专区|
99久久免费精品国产72精品九九
|
亚洲成a人片77777群色|
中文字幕无码精品亚洲资源网|
麻豆成人久久精品二区三区免费|
色婷婷五月综合亚洲小说|
亚洲另类自拍丝袜第五页|
午夜三级a三级三点在线观看|
人人综合亚洲无线码另类
|
曰本无码人妻丰满熟妇5g影院
|
激情内射亚州一区二区三区爱妻|
欧美人与禽交zozo|
精品无码日韩国产不卡AV|
综合图区亚洲欧美另类图片|
日本熟妇厨房xxxxx乱|
日韩免费视频一一二区|
欧美最猛性xxxx|
国产又黄又爽胸又大免费视频|
亚洲午夜成人精品无码APP|
成人做爰黄AAA片免费看少妃|
99久久亚洲精品无码毛片|
小说 亚洲 无码 精品|
免费高清理伦片a片在线观看
|
亚洲精品在看在线观看|
精品久久国产综合婷婷五月|
国产乱子轮XXX农村|
国产成人精品亚洲午夜|
亚洲综合色婷婷七月丁香|
日韩精品一区二区三区中文|
欲色影视天天一区二区三区色香欲
|
无套内射chinesehd|
国产成人精品无缓存在线播放
|
亚洲午夜福利精品久久|
日本少妇BBW丰满做爰图片|
麻花传媒MV一二三区别在哪里看|
欧美老熟妇xb水多毛多|
麻豆国产人妻欲求不满|
《艳香艳史》在线播放|
麻豆潘甜甜传媒77777|
乱japanese偷窥piss|
另类内射国产在线|
奇米网777色在线精品|
50岁熟妇高潮惨叫|
中文字幕无码AV波多野吉衣|
漂亮人妻被黑人久久精品|
一本大道东京热无码|
久久久久99精品成人片牛牛影视|
精精国产xxxx视频在线|
亚洲爆乳精品无码一区二区三区|
久久96国产精品久久久|
在线精品无码AV不卡顿|
三亚在线观看免费高清电视剧软件|
日韩人妻无码一区二区三区综合|
在线观看免费的成年影片|
亚洲欲色欲色XXXXX在线观看|
骚虎av在线网站|
人妻丰满熟妇AV无码片|
人妻人人添人妻人人爱|
欧美专区日韩视频人妻|
欧美巨大巨粗黑人性aaaaaa|
亚洲精品成人区在线观看|
成人性生交A片免费看武则天|
欧美FREESEX8一10精品|
免费岛国av动作片在线观看|
欧美乱大交xxxxx潮喷|
亚洲欧美日韩另类丝袜一区|
国产农村妇女毛片精品久久|
国产精品永久免费|
亚洲一区二区三区丝袜|
不戴套挺进朋友人妻|
国产人久久人人人人爽|
国第一产在线无码精品区|
免费 人妻 无码 不卡中文...|
国产sm重味一区二区三区|
亚洲gv白嫩小受在线观看|
精品综合久久久久久888蜜芽|
99成人做爰A片免费看网站
|
人妻 自慰 偷窥 无码|
亚洲综合色自拍一区|
亚洲国产精品日韩AV专区|
热思思99RE久久精品国产首页|
久久久久久久极品内射|
一夲道中文字幕av高清片|
成人午夜又粗又硬又长|
亚洲综合色视频在线观看|
精品无码久久久久成人漫画|
国产一区二区三区免费播放|
最新69国产成人精品视频免费|
精品国产午夜福利在线观看|
中文字幕人成人乱码亚洲电影
|
日本道色综合久久影院|
孕妇特级毛片ww无码内射|
国产在线乱码一区二区三区|
77777亚洲午夜久久多喷|
欧美不卡一区二区三区|
国产精品无码无片在线观看3D|
色综合久久中文综合网|
超碰97久久国产精品牛牛|
婷婷五月综合缴情在线视频|
精品久久久久久久久久久aⅤ|
少妇人妻精品一区二区三区|
国产精品无码专区在线播放|
天天综合网久久综合免费人成
|
2022久久国产精品免费热麻豆|
少妇被粗大的猛烈进出VA视频|
国产一区二区三区不卡在线观看|
国产精品视频一区国模私拍|
国产亚洲精品线观看动态图|
男女裸体做爰视频免费|
国产精品毛片一区二区|
国产粉嫩高中无套进入|
先锋av资源|
日本理伦少妇4做爰|
国产a∨国片精品白丝jk制服|
国产精品午夜小视频观看|
亚洲国产精品无码久久久秋霞1|
香蕉久久AV一区二区三区|
99草草国产熟女视频在线|
美国人XXXXXL19|
国语自产精品视频在 视频|
日韩经典精品无码一区|
国产亚洲情侣一区二区无码av|
少妇无码av无码专区在线观看|
蜜桃无码一区二区三区|
精品剧情v国产在线观看|
国产丝袜视频一区二区三区
|
在线观看高清黄网站免费|
精品国产亚洲一区二区三区在线观看|
无码人妻A片一区二区三区不卡
|
日韩国产综合精选|
国产精品福利自产拍在线观看|
性饥渴艳妇性色生活片在线播放|
中文无码av在线亚洲电影|
国产精品多人P群无码|
久久超碰97人人做人人爱|
亚洲成av人片一区二区梦乃|
日本午夜精品一区二区三区电影|
国产av巨作情欲放纵|
精品午夜国产福利观看|
中文字幕av无码一区二区三区电影
|
国产一极内射視颍一|
强被迫伦轩高潮无BD|
久久免费99精品国产自在现线|
国产免费踩踏调教视频|
青青青国产依人在线|
亚洲免费精品AⅤ国产|
熟妇人妻无乱码中文字幕真矢织江
|
性色av无码不卡中文字幕|
樱花草视频www日本韩国|
边吃奶边狠狠躁日韩A片|
亲子乱aⅴ一区二区三区下载|
久久久无码精品亚洲A片猫咪|
国产精品一区二区久久精品|
成在线人免费视频|
激情欧美日韩一区二区|
日韩亚洲国产中文字幕欧美
|
欧美熟妇丰满XXXXX裸体艺术|
亚洲高清乱码午夜电影网|
无码精品人妻一区二区三区AV|
少妇被爽到高潮动态图|
精品一区二区三区无码AV久久
|
国产亚洲精品aaaa片小说|
2021国产精品午夜久久|
色妺妺视频网|
国产最新AV在线播放不卡|
精品香蕉一区二区三区|
中日韩精品视频在线观看|
网友自拍区视频精品|
69久久精品无码一区二区|
亚洲日本VA中文字幕|
日本人妻人人人澡人人爽|
国产精品视频色尤物yw|
精品人妻无码一区二区三区不卡|
亚洲日韩成人无码不卡网站|
国产强被迫伦姧在线观看无码|
国产三级精品三级在线专1|
无码三级香港经典三级在线视频|
午夜成人无码福利免费视频|
国产成年无码av片在线|
99久久精品费精品国产一区二|
国产18禁黄网站免费观看|
一线高清中文免费观看视频|
麻豆精品偷拍人妻在线网址|
人妻少妇中文字幕乱码|
欧美熟妇丰满XXXXX裸体艺术|
宅男午夜成年影视在线观看|
在线观看日本亚洲一区|
人妻夜夜爽爽88888视频|
国产精品自在线拍国产手青青机版
|
麻花豆精产三产区区别|
FREEZEFRANE丰满老师|
亚洲国产欧美在线人成AAAA|
天堂无码人妻精品av一区|
国产成人A亚洲精V品无码|
最近在线更新8中文字幕免费|
国产精品午夜自在在线|
成在人线av无码免费看网站直播|
在线亚洲精品国产二区图片欧美|
人妻熟妇乱又伦精品视频|
最近最新中文字幕|
日本午夜精品一区二区三区电影|
少妇人妻偷人精品视频|
亚洲精品无码不卡久久久久|
亚洲精品久久久久|
久久人人爽人人爽人人av|
呦系列视频一区二区三区|
无码人妻精品一区二区在线视频
|
少妇无码av无码一区|
少妇精品无码一区二区三区|
亚洲欧美日韩国产制服另类|
亚洲精品久久|
老司机午夜福利视频|
男女吻摸下面一进一出视频|
一女被二男吃奶a片免费观看|
国产精品久久久国产盗摄|
免费国产A国产片高清|
国产激情视频在线观看的|
婷婷五月花|
亚洲 自拍 另类小说综合图区|
丰满人妻一区二区三区免费视频|
内射国产内射夫妻免费频道|
不戴套挺进朋友人妻|
东京热无码人妻一区二区AV|
久久精品成人免费观看三|
亚洲成av人片无码天堂下载|
国产在线观看无码的免费网站
|
亚洲日产无码中文字幕|
成人网站国产在线视频内射视频|
印度女人狂野牲交|
女厕厕露P撒尿八个少妇|
狠狠色综合激情丁香五月|
国产亚洲精品A第一页|
精品一区二区三区影院在线午夜|
天堂中文www官网天堂最新版|
久久久亚洲欧洲日产国码二区
|
日本中文字幕在线|
国产suv精品一区二区五|
国内精品久久人妻无码妲己影院|
亚洲国产成人精品无码区在线|
毛片内射久久久一区|
亚洲乱码AV中文一区二区|
国产成人精品久久一区二区|
欧美xxxxx精品|
yy1111111少妇影院光屁股|
久久精品人妻无码专区|
麻豆成人久久精品二区三区免费|
亚洲精品无码久久久久|
人妻少妇无码中文幕久久|
久久AV国产麻豆HD真实乱|
中文字幕一区二区三区精华液
|
日本人妻人人人澡人人爽|
全球AV集中精品导航福利|
国产精品视频一区二区三区无码|
韩国精品久久久久久无码|
亚洲精品偷拍av一区二区|
久久婷婷五月综合色中文字幕|
五月综合网亚洲乱妇久久|
乱人伦人成品精国产在线|
国产高清一区二区三区视频|
单身男女高清免费观看视频|
国产色情三级A区色情男女
|
国产精品无码无片在线观看|
《人妻》波多野结衣|
国产又A又黄又潮娇喘视频|
国产成熟女人性满足视频|
精品国产精品久久一区免费式|
野狼AV午夜福利在线|
国产瑜伽白皙一区二区|
大地影院mv高清在线观看免费
|
色综合视频一区中文字幕|
国产精品无码专区在线播放|
在线观看无码AV网址|
国产精品成熟老女人|
永久免费无码AV网站在线观看|
午夜性刺激免费看视频|
久久婷婷五月综合国产尤物APP|
黑寡妇巨大粗爽毛片欧美|
亚洲 国产 韩国 欧美 在线|
亚洲午夜成人精品无码色欲|
久久久综合九色合综|
少妇伦子伦精品无吗|
色欲人妻AAAAAAA无码|
国产男女猛烈无遮挡免费网站|
免费福利视频一区二区三区高清
|
人妻精品动漫h无码专区|
美女裸体无遮挡免费视频网站|
白又丰满大屁股bbbbb|
少妇高潮无套内谢麻豆传|
欧美日韩亚洲国产精品|
久久亚洲色WWW成人欧美|
CHINESE性内射高清国产|
999久久国产精品免费人妻|
国产亚洲日韩在线三区|
麻花星空MV高清免费|
欧美xxxxxbbbb|
午夜精品久久久久久久久久久久|
人妻少妇久久中文字幕|
给我免费播放片高清在线观看视频|
国产精品久久久久久久久久久久午衣片
|
国产午夜亚洲精品不卡|
国产网战无遮挡|
国产精品一区二区国产主播
|
亚洲综合精品成人|
欧美性狂猛xxxxxbbbbb|
性大毛片视频|
大肉大捧一进一出好爽mba|
玩弄了裸睡少妇人妻野战|
国产情侣久久久久aⅴ免费|
亚洲成AV人无码中文字幕|
精品人妻一区二区三区浪潮在线|
日本特黄无码毛片在线看|
国产成人精品免费视频网页大全
|
国产精品无码无片在线观看3D
|
亚洲欧美日韩愉拍自拍美利坚|
国内老熟妇对白HDXXXX|
又色又爽又黄18禁美女裸身无遮挡|
久久久精品免费|
中文字幕精品久久久久人妻红杏1|
久久精品中文字幕一区二区三区|
欧美性生恔XXXXXDDDD|
国产午夜精品一区二区三区|
麻豆国产尤物av尤物在线看
|
黑人啊灬啊灬啊灬快灬深|
亚洲国产成人手机在线电影
|
日本熟妇厨房xxxxx乱|
精品久久久久久亚洲综合网|
无码喷水一区二区浪潮av|
免费看成人羞羞视频网站在线看|
中文字字幕在线中文乱码|
亚洲国产成人精品女人久久久|
国产目拍亚洲精品二区|
男女啪啪免费体验区|
在线看片免费观看视频|
无套内谢老熟女|
日韩亚洲国产主播在线不卡|
亚洲最新无码成AV人|
成人性生交大免费看|
亚洲欧美日韩国产制服另类|
亚洲成av人片在线观看|
国精品产区WNW2544|
日产国产亚洲a片无码app下载|
亚洲人成精品久久久久|
98精产国品一二三产区区
|
久久久久久欧美精品无码超碰|
人妻丰满熟妇av无码区app|
国产美女a做受大片观看|
天天摸夜夜添夜夜无码|
最近日本mv字幕免费高清在线|
欧美三级A做爰在线观看|
免费看国产成年无码av|
又湿又黄裸乳漫画无遮挡网站|
国产亚洲欧美日韩一区图片|
三级4级做a爰60分钟|
99久re热视频这只有精品6|
欧美伦理毛片在线看|
成人做爰A片免费看网站性晶|
天天摸夜夜添夜夜添A片小说|
欧美性猛交xxxx黑人|
无码视频一区二区三区|
铜铜铜铜铜铜铜铜好大好深色|
一本大道熟女人妻中文字幕在线|
亚洲男人的天堂在线播放|
国产精品永久久久久久久久久|
无码人妻AⅤ一区 二区 三区
|
久久精品国产精品亚洲蜜月|
亚洲日韩在线a在线观看|
日韩一卡2卡3卡4卡新区亚洲|
久久99久久99精品免视看|
已满十八岁从此入戴耳机的意思|
另类亚洲欧美精品久久|
中文字幕无码第1页|
青春草在线视频免费观看|
99久久无色码中文字幕|
国产亚洲情侣一区二区无码av|
激情综合色五月丁香六月亚洲|
亚洲色大成网站www|
亚洲女同成av人片在线观看|
午夜福利92国语|
亚洲精品国产精品国自产观看|
韩国v欧美v亚洲v日本v|
熟妇女人妻丰满少妇中文字幕|
欧美高清性色生活片免费观看|
熟妇人妻中文字幕|
欧美又粗又大AAA片|




能檢查出什么,白帶常規(guī)檢查的注意事項(xiàng).png)
氧運(yùn)動(dòng)是什么,有哪些好處.png)
時(shí)驚厥的癥狀,如何預(yù)防小兒驚厥.png)

痛如何才能緩解一文科普<div id=)




 、調(diào)情志
、調(diào)情志